Development of new tools for plant genetic engineering
While the basic strategy for assembly of multigene construct is now well established and functional, we are expanding the MoClo system by creating novel components including new vectors and parts. One example of a project consists of the development of promoter libraries to co-express several genes of interest in the same cell type of a plant. With this system (developed in collaboration with the group of Alain Tissier), synthetic promoters were made consisting of a binding site for a synthetic designer TAL effector (a 19 bp sequence) flanked by random sequence. These synthetic TAL activated promoters (STAPs) become active only when bound to a corresponding designer TAL effector. Multigene constructs are then made containing the genes that need to be co-expressed under the control of synthetic promoters, plus a designer TAL effector placed for example under control of a tissue-specific promoter (Fig. 2). Using this strategy, all genes placed under control of a STAP will be co-expressed in the same cells of a plant in which the designer TAL effector will be expressed, for example in petals or pollen (Fig. 2).
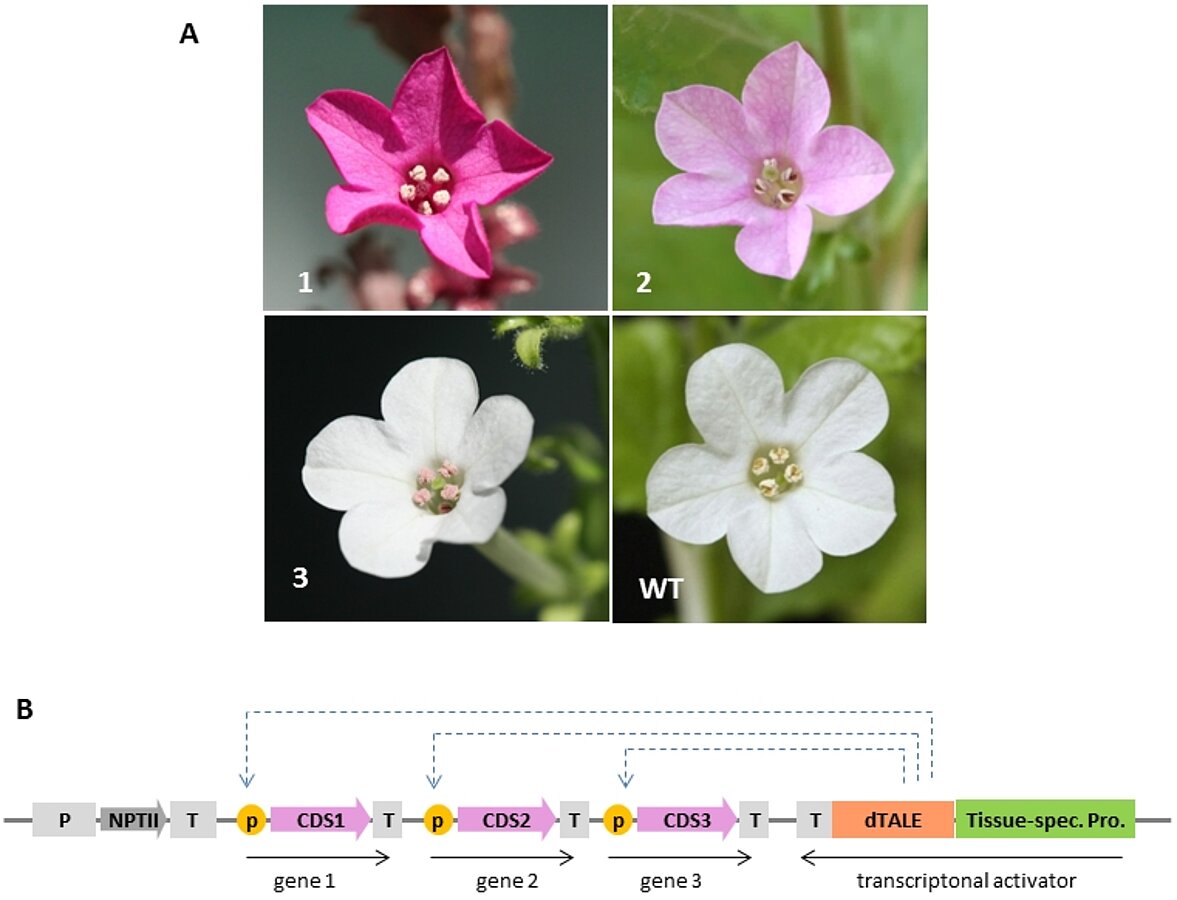
Fig. 2: Tissue-specific expression of a synthetic betanin biosynthesis pathway in Nicotiana benthamiana. (A) Flowers of wild type and transgenic Nicotiana benthamiana plants expressing three genes required for betanin biosynthesis in many tissues (1), petals (2) or pollen (3). (B) Structure of the constructs used for making the transgenic plants showed in (A). The TAL effector-based transcriptional activator binds to sequences present in 3 synthetic TAL activated promoters (p) resulting in transcription of the 3 genes of the pathway in cells where the tissue-specific promoter is active.
Tools for gene editing in plants
The elucidation of the mode of action of CRISPR in 2012 has led to a revolution in gene editing that allows researchers to alter any sequence of interest in any organism of choice, including plants. Nevertheless, the efficiency of gene editing can vary widely in experiments, depending on the Cas9 gene that is used and the particular target that one desires to modify. We have optimized the sequence of the Cas9 nuclease for high level expression in Arabidopsis. Constructs containing the optimized Cas9 sequence and carrying guide RNA sequences designed to target two genes involved in trichome development led to the generation of knock out (hairy) phenotypes in more than 70% of primary Arabidopsis transformants (Fig. 3). This Cas9 gene will be tested with other target sequences and in other plant species.

Fig. 3: Arabidopsis primary transformants with an optimized Cas9 construct targeting 2 genes involved in trichome development (A). More than 70% of the transformants have a hairy phenotype, indicating that the 4 alleles of both genes have been knocked out. (B), Plant obtained with a control construct with non-optimized Cas9.
The Newcotiana project (https://newcotiana.org/), a project financed by the European Commission, aims to generate Nicotiana benthamiana and tobacco lines for production of substances with added value used in medicine and cosmetics. The Nicotiana lines will be generated without using standard genetic engineering tools to avoid making transgenic plants. Instead, they will be made using New Plant Breeding (NPB) methods that rely on the use of cis-genic sequences that are derived either from the host plant or from plants with which it can hybridize. For this project, we are developing NPB tools, including nucleases that will not integrate in the Nicotiana genome. We will also generate a library of cis-genic parts for use in Nicotiana benthamiana and tobacco.
CRISPR gene editing usually leads to a few nucleotide deletions or insertions in a target gene, and usually results in inactivation of this gene. In contrast, gene-targeting aims to insert complete functional genes at a chosen chromosomal position. In the GeneReplace project, a BMBF-funded project made in collaboration with Prof. Alain Tissier and Prof. Bettina Hause, we are developing protocols for improving the efficiency of gene targeting in tomato. This project aims to increase the frequency at which gene-targeting can occur at a desired locus, but also aims to improve the efficiency of plant regeneration in tissue culture, which is still a bottleneck for tomato transformation in many cultivars.
Development of parts and vectors for expression in yeast
Yeast genetic engineering can be done by introducing genetic material either on one of the yeast chromosomes or on extrachromosomal replicating episomes. We are developing vectors and parts for engineering yeast using both approaches.
One of the advantages of the MoClo system is to allow production of libraries of constructs rather than only individual constructs. As an example, a library of constructs for beta-carotene biosynthesis was made, where the promoter and terminators of each construct were randomly cloned from libraries of 10 promoters and 10 terminators (Fig. 4). Therefore, no two constructs from this library are expected to be identical. The best-working promoter/terminator combination can be screened visually by the orange color of beta-carotene in this case. We are also developing vectors and parts for integrating constructs on the yeast chromosome using the CRISPR/Cas9 system.
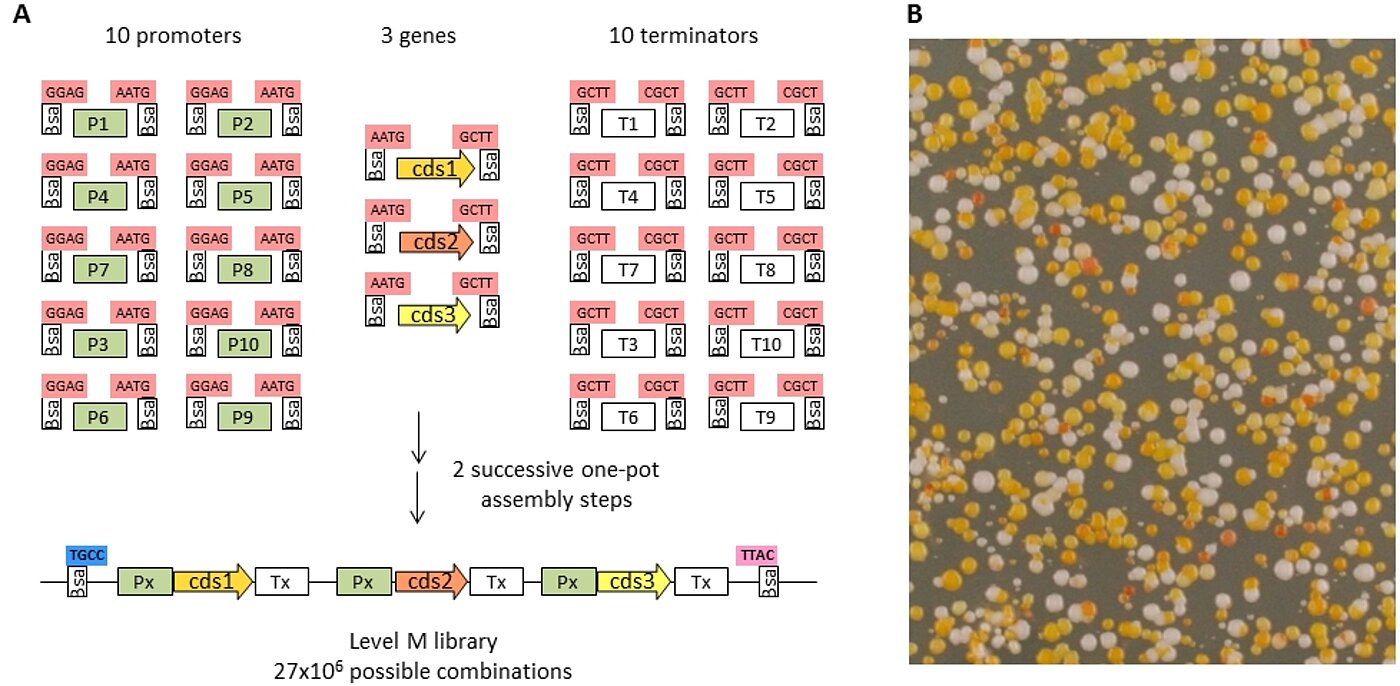
Fig 4: Construction of a library of carotenoid producing yeast cells. A: A library of constructs is made using two one-pot assembly steps starting from 2 libraries of 10 promoters and 10 terminators and 3 genes required for biosynthesis of beta-carotene. The library prepared using E. coli cells is then transformed into yeast cells. B: library of carotenoid constructs transformed into yeast cells.
Engineering of plant biosynthetic pathways

Fig. 5: Fruit of Bixa orellana with seeds. The seeds are covered with a red substance containing the apocarotenoid bixin. Photo: M.H. Walter.
We are interested in plant biosynthetic pathways responsible for production of colorful metabolites such as betalains, anthocyanins and carotenoids. These pathways can be used as model pathways for toolbox development, but can also have biotechnological applications. For example, betalains are used as natural food colorants and are thought to have health-promoting properties. However, their restricted distribution in plants of the caryophyllales order has the consequence that betalains are not naturally present in the great majority of commonly consumed plant products. Therefore, we are introducing the entire pathway in plants that do not normally make betalains, such as tomato.
Moreover, we also try to elucidate missing genes/enzymes in pathways, whose description is still incomplete. For example, one carotenoid derivative of interest is bixin, which is used industrially as a natural food colorant. Bixin naturally accumulates in the seeds of the tree Bixa orellana (Fig. 5) and is thought to be produced by enzymatic cleavage and modification of a carotenoid precursor. We are now looking for some of the still elusive genes coding for enzymes of the bixin pathway for engineering the whole pathway in heterologous hosts.
This page was last modified on 27 Jan 2025 27 Jan 2025 27 Jan 2025 27 Jan 2025 27 Jan 2025 04 Feb 2025 .