MAMP-induced calcium signaling
A rapid MAMP-induced rise in cytosolic [Ca2+] is an early signalling event essential for downstream signal transduction. We adapted the apoaequorin-based [Ca2+] monitoring system into a high-throughput microplate-based assay (Fig. 1) for genetic screens. Mutants with a “changed calcium elevation” (cce) response to the bacterial MAMP, flg22, were isolated and revealed novel mutant alleles of receptor complex components (receptor, co-receptor and associated proteins; Ranf et al., 2011, 2012, 2014) and the membrane protein quality control pathway (i.e. controlling protein/receptor glycosylation; Trempel et al., 2016). The predominance of receptor mutants among the “MAMP-specific” cce mutants demonstrated that such a screen is well suited for finding the receptors of “orphan MAMPs”. Indeed, as proof-of-principle, we identified the first putative plant receptor for bacterial lipopolysaccharide-derived MAMP (Ranf et al., 2015). With this approach, we aim to identify additional receptors and signaling components and elucidate their roles in signal processing during plant-microbe interactions.
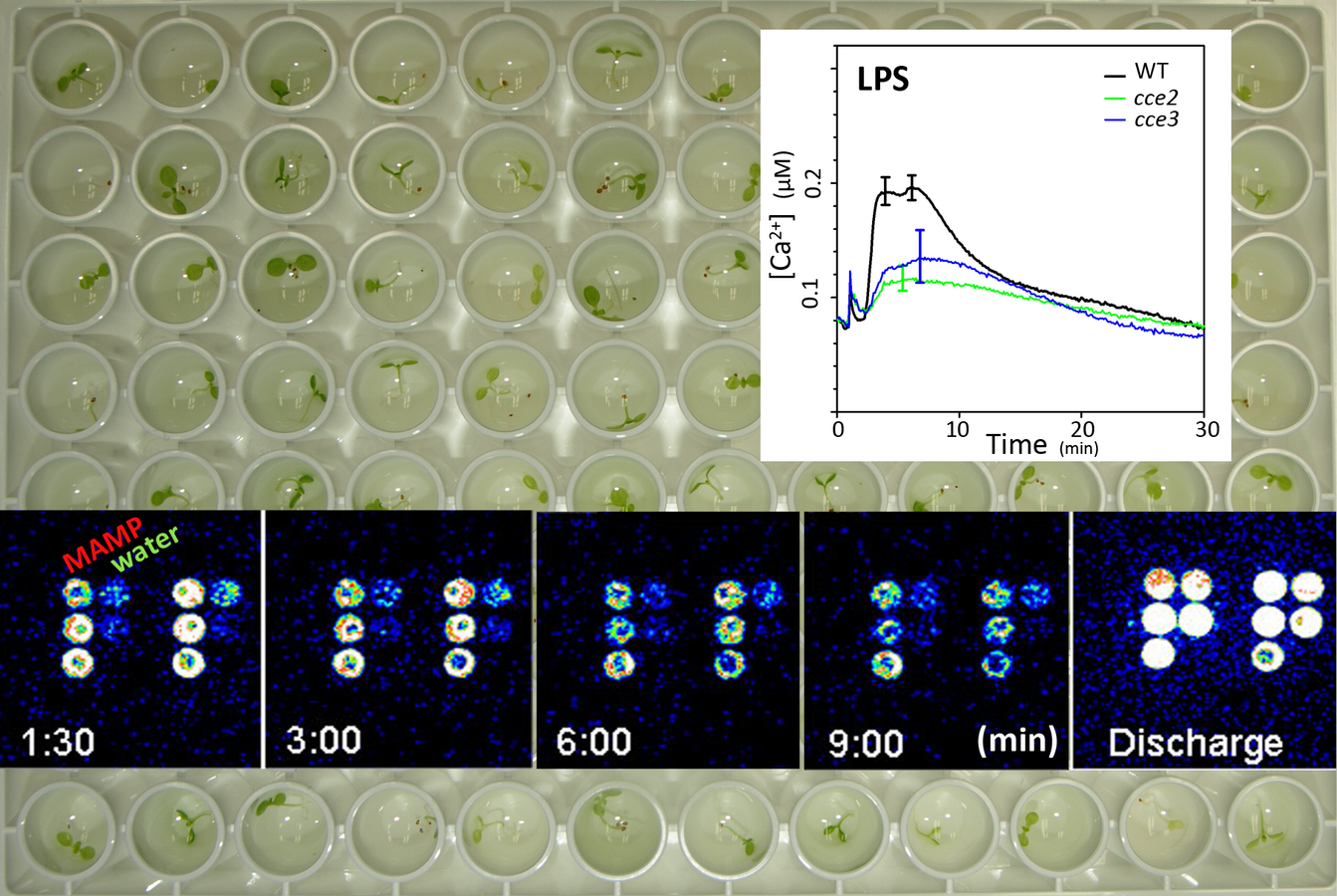
Fig. 1: MAMP-induced changes in cytosolic [Ca2+] can be visualized as heightened luminescence in Arabidopsis plants expressing the Ca2+ reporter, apoaequorin. This microplate-based assay is highly amenable for high throughput genetic mutant screens. Inserted graph shows the Ca2+ response to bacterial lipopolysaccharide (LPS) in two “changed calcium elevation” (cce) mutants.
Control of plant defense via mitogen-activated protein kinases (MAPKs)
MAPKs are important control elements of plant defense (Lassowskat et al., 2014, Lee et al., 2015) but the same MAPK components often also regulate other cellular pathways. The preservation of signal fidelity without erroneous crosstalk between pathways is still not well understood in plants but involves multiple mechanisms such as gene expression regulation of the MAPK elements, the MAPK substrate diversity, as well as the presence of pathway-specific protein complexes. We focus on identifying MAPK-interacting proteins and MAPK substrates, and elucidating the impact of phosphorylation on their function. As activated MAPKs shuttle into the nucleus to regulate transcriptional processes, emphasis is placed on transcription factors and co-regulators (Bethke et al., 2009, Pecher et al., 2014) but additionally, we are also interested in MAPK-mediated regulation of post-transcriptional processes in plant defense (Maldonado-Bonilla et al., 2014, Tabassum et al., 2019), secretion-related processes (e.g. Exo70, Teh et al., 2018) and phospholipid signaling (Hempel et al., 2017, Menzel et al., 2019). Different strategies are being pursued to isolate MAPK substrates and interacting proteins; this includes yeast-two hybrid screens, mass-spectrometry-based identification of interactors, phosphoproteomics and protein array-based screens for MAPK substrates (Fig. 2).
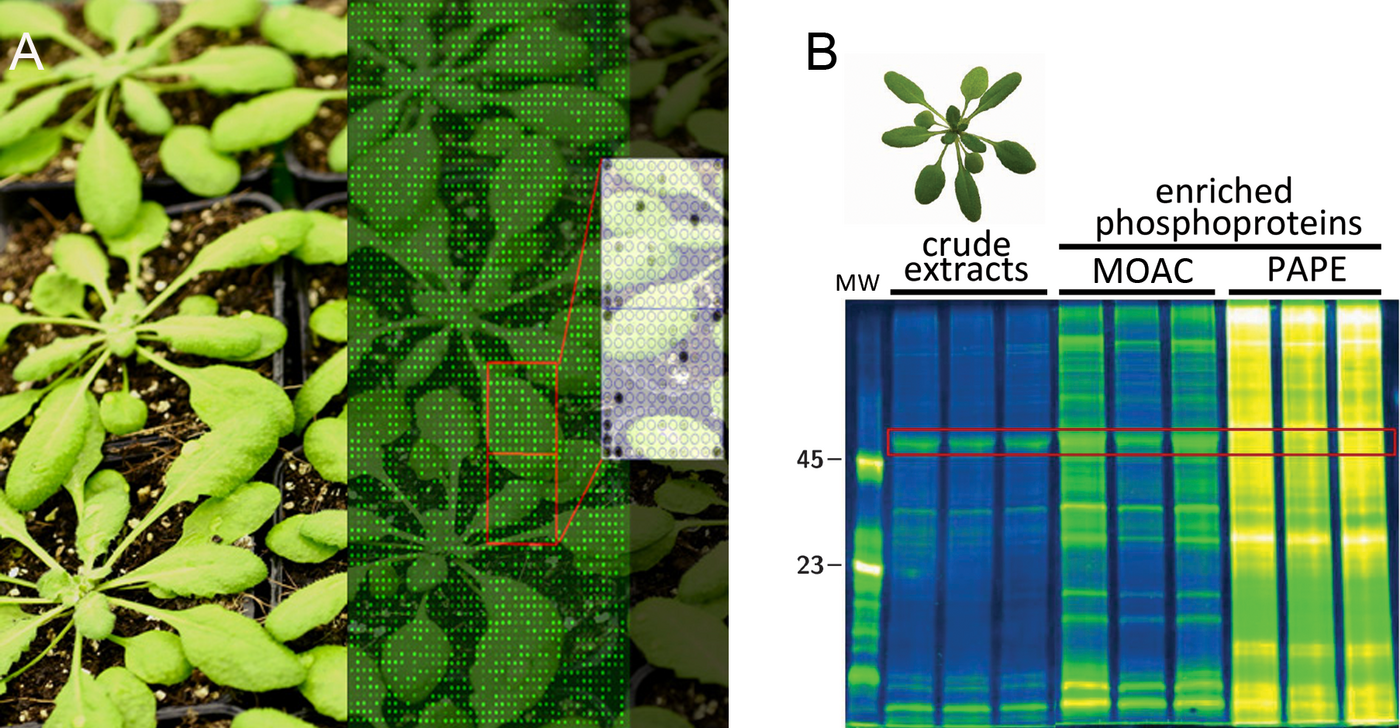
Fig. 2: Examples of methods used for identifying MAPK targets: (A) An Arabidopsis protein array used for in vitro screening of MAPK substrates (Image superimposed on Arabidopsis plants). (B) Phosphoprotein-staining (ProQ Diamond) gel comparing phosphoproteins enriched using classical MOAC (Metal-Oxide-Affinity Chromatography) and the improved PAPE (Prefractionation-Assisted Phosphoprotein Enrichment) method (Lassowskat et al., 2013).
Suppression of defense by pathogen effectors
Bacterial pathogens such as Pseudomonas syringae or Xanthomonas campestris use a specialized so-called type III secretion system to inject effectors into plant cells (Fig 3), which interfere with defense signaling (e.g. XopH, Blüher et al., 2017; HopQ, Zembek et al., 2018). The well-studied P. syringae effector, AvrRpt2 (and several bacterial homologs), blocks specifically one of the two branches of flg22-responsive MAPKs (Eschen-Lippold et al., 2016). Such a selective interference of MAPKs presumably represents a pathogen strategy to fine-tune MAPK-regulated cellular signaling. This function of the effector is independent of RIN4 (an important AvrRpt2 target regulating plant defense) but requires its cysteine protease activity, which may therefore trigger also the N-end rule protein degradation pathway (Goslin et al., 2019). We aim to identify the additional AvrRpt2 target(s) responsible for this inhibitory effect on MPK4/MPK11 activation and uncover the impact on defense signaling and disease resistance.

Fig. 3: (A) Electron micrograph showing flagella of Pseudomonas syringae pv tomato DC3000 (Magnification: 31 500-fold), a source of the flg22 MAMP peptide that elicits defense responses. (B) Besides the flagella, thinner pili structures (6-8 nm in diameter; highlighted by arrow) are seen, which act as conduits for effector proteins into plant cells (Magnification: 63 000-fold). (C) Scheme illustrating the injection of the AvrRpt2 effector protein into a plant cell and the interference of one of the two known branches of MAPK cascades that are activated by flagella-derived flg22.
This page was last modified on 27 Jan 2025 27 Jan 2025 27 Jan 2025 27 Jan 2025 .